![]()
Lo que no te mata te hace más… resistente
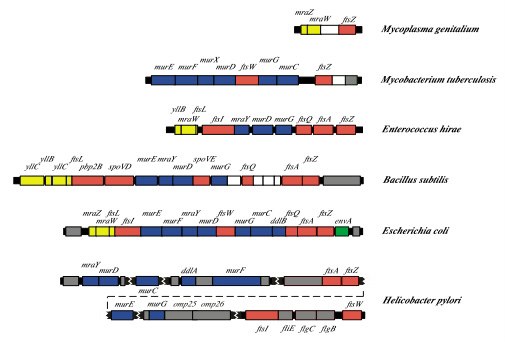
Hace unos meses comentábamos aquí que con los antibióticos a veces menos es más. Pero ¿qué ocurre cuando no sólo eso no es posible, sino que el paciente necesita un tratamiento antibiótico muy largo? Pues parece que no pasa gran cosa y que lo que no te mata te hace más… resistente.